
June 26, 2026 · 12:24 AM
Area MT/V5: How the Brain Sees Motion
Area MT/V5 shows how visual cortex turns noisy moving dots into a usable motion signal, and why lesion, recording, and microstimulation studies made it a model system for linking neurons to perception.

You notice MT most when it fails quietly. A glass of water is easy to see on a table, but to reach for it you also need to know that your hand is moving, the rim is drifting across your retina, and the world is not sliding sideways. Area MT, also called V5 in humans, is one of the brain's best-studied answers to that problem: it is a patch of extrastriate visual cortex where many neurons are tuned for the direction and speed of motion.
MIT's Human Brain sequence uses MT as a worked example of how cognitive neuroscience links a mental ability to a specific cortical region: first identify a candidate area, then ask whether its activity is merely correlated with a percept or actually helps cause it. Nancy Kanwisher's course page places "Visual Motion Area MT" immediately after the early V1 mapping topics, which is exactly where it belongs in the learning sequence: after retinotopy, orientation, ocular dominance, receptive fields, and cortical magnification, MT shows how visual cortex begins to build a more behaviorally useful representation. 1
The concept: a motion-specialized visual area
Area MT/V5 is not a tiny movie screen. It is a cortical field, downstream from early visual cortex, whose neurons respond strongly to moving visual patterns and often prefer a particular direction and speed. Reviews describe MT as a historically important model for cortical processing streams, specialized visual representations, and the relationship between neural activity and perception. 2
The core intuition is simple: V1 contains many neurons that care about local edges and local motion, but real motion perception usually requires integrating many noisy local samples. A random-dot display, for example, may contain hundreds of dots, only some of which share a common direction. MT is one place where those local signals are pooled into a stronger estimate of "which way the pattern is moving."
Anatomically, macaque MT lies in extrastriate cortex near the superior temporal sulcus; human work often refers to the homologous motion-responsive complex as MT+ or V5/MT. The naming is a reminder that cross-species mapping is not perfect. "MT" is the classic monkey area; "V5" and "MT+" are common human imaging terms for a motion-responsive region or cluster.
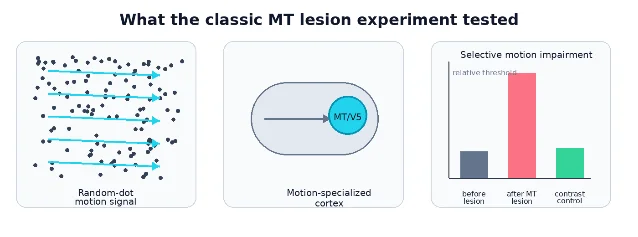
The landmark evidence: remove MT, and motion perception suffers
The landmark paper for this concept is Newsome and Paré's 1988 Journal of Neuroscience lesion study. They trained rhesus monkeys on two tasks: a motion task using dynamic random-dot displays, and a contrast task using stationary gratings. Then they injected ibotenic acid into MT and measured what changed. The result was selective: motion thresholds rose strikingly, while contrast thresholds showed little or no impairment. 3
That result mattered because it moved MT beyond "interesting physiology." Many regions respond to a stimulus. Fewer pass the stronger test: when the region is disrupted, the perceptual ability gets worse. The Newsome and Paré experiment made MT a causal candidate for motion perception, not just a place where motion-selective cells happened to sit.
The stimulus design was also important. Random-dot motion lets the experimenter vary motion coherence: how many dots move together in one direction while the rest behave as noise. That gives a graded psychophysical threshold rather than a yes/no observation. It also gives neuroscientists a way to compare behavior with neural responses under tightly matched conditions.
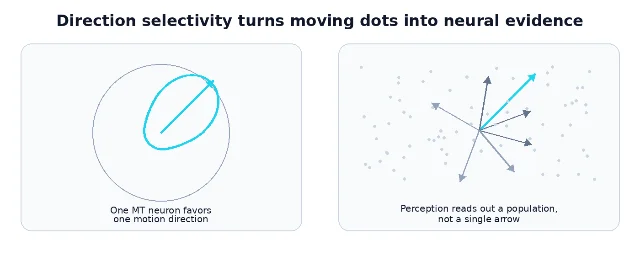
What MT neurons encode
Single-unit recordings helped define the computational personality of MT. Maunsell and Van Essen recorded from 168 histologically identified MT units in macaques and reported a high degree of direction selectivity, strong speed tuning, and preferred speeds spanning roughly 2 to 256 degrees per second. 4 Albright's 1984 comparison of macaque MT and V1 found that all 110 MT neurons studied were directionally selective for one or more moving stimuli, supporting the idea that MT is a further specialization over V1 for motion processing. 5
That does not mean every MT neuron is a pure "motion detector" in the cartoon sense. MT neurons have receptive fields, preferred directions, preferred speeds, tuning bandwidths, and context-dependent responses. Some respond differently to component motion and pattern motion; some carry disparity information; attention and task demands can modulate activity. The useful simplification is not "MT equals motion." It is: MT is one of the strongest cortical examples of a population code for visual motion.
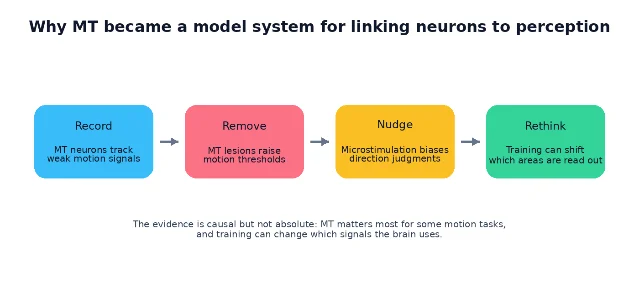
From correlation to causation: record, remove, nudge
MT became famous because researchers could connect three kinds of evidence in the same conceptual system.
First, recording studies showed that MT neurons carry signals strong enough to explain behavioral discrimination. Britten, Shadlen, Newsome, and Movshon compared monkeys' psychophysical performance with single-neuron responses during matched random-dot tasks. Under those conditions, many MT neurons had sensitivity similar to the animal's own motion-discrimination sensitivity. 6
Second, lesion and inactivation studies showed necessity under particular task conditions. Newsome and Paré's selective impairment result is the cleanest introductory example: disrupt MT, and motion thresholds rise. 3
Third, microstimulation studies showed that adding a small artificial signal to MT can bias what the animal reports seeing. In the 1990 Nature experiment, Salzman, Britten, and Newsome stimulated direction-selective MT neurons while monkeys judged motion direction; the stimulation biased choices toward the neurons' preferred direction. 7 A larger 1992 Journal of Neuroscience report found statistically significant effects in 89 experiments; in 86 of those 89, stimulation pushed judgments toward the stimulated neurons' preferred direction. 8
That triplet is why MT is so useful pedagogically. Recording says "the signal is there." Lesion says "the signal can be necessary." Microstimulation says "changing the signal can change perception."
Human MT and the strange case of motion blindness
Human evidence came from several directions. Brain-imaging studies found motion-responsive regions near the expected lateral occipito-temporal territory; Orban and colleagues localized a human area involved in processing contours defined by motion differences, linking human imaging to the V5/MT tradition. 10
Clinical neuropsychology offered a more vivid clue. Zihl, von Cramon, and Mai described a patient with bilateral posterior brain damage who had a relatively selective disturbance of visual movement perception. She lacked normal impressions of movement in depth, had no visual motion aftereffects or apparent phi movement, and could perceive central motion only when target velocity stayed below 10 degrees per second. The authors concluded that movement vision depends on mechanisms beyond primary visual cortex, involving lateral temporo-occipital cortex and underlying white matter. 11
This condition is often called akinetopsia, or motion blindness. It is rare, and it should not be reduced to a simple "MT is broken" slogan. Still, it gives a powerful phenomenological hint: motion is not merely the same visual scene updated over time. The brain has specialized mechanisms for seeing motion as motion.
What MT does not mean
MT is a success story, but the success can mislead if stated too strongly.
First, MT is not the only motion area. Human PET and fMRI studies detect broader motion-responsive networks, and monkey visual cortex contains multiple motion-related regions, including MST and parietal areas. The right statement is not "motion lives in MT." It is "MT is a central hub in a wider motion-processing system."
Second, MT is not always necessary in the same way. Liu and Pack reported that MT's causal contribution to motion discrimination can depend on training history: inactivation can severely impair discrimination after some training regimes, but have little detectable influence after others. Their interpretation is that training can shift the readout of motion information between MT and lower-level cortical areas. 9
That finding is not an embarrassment for the classic account. It is an upgrade. It says that a brain region can be highly informative and causally powerful without being the only possible route to a percept. The brain may learn which signals to trust.
Why this concept matters
MT/V5 teaches three durable lessons about cognitive neuroscience.
One region can be functionally specialized without being isolated. MT has a recognizable computational bias: direction and speed of visual motion. But it sits inside a network of inputs, outputs, task demands, and learned readout strategies.
Causal evidence comes in degrees. A beautiful activation map is useful, but MT became a model system because scientists combined recording, lesion/inactivation, behavior, and stimulation. That is the method lesson students should carry into later topics such as face areas, place areas, language regions, and theory-of-mind networks.
Perception is an inference from noisy signals. Random-dot motion tasks make this visible. The stimulus contains ambiguity; the animal or person must decide whether weak evidence supports one direction over another. MT helped turn perception from a vague mental capacity into a measurable neural decision problem.
In that sense, MT is more than a motion area. It is a bridge between early visual coding and the central question of cognitive neuroscience: how patterns of neural activity become the contents of experience.
Landmark paper: Newsome & Paré, 1988, "A selective impairment of motion perception following lesions of the middle temporal visual area (MT)".
Course connection: MIT/Nancy's Brain Talks 3.9, "Example: Visual Motion Area MT" follows the early visual cortex topics and shows how MT turns visual-area mapping into a causal test of perception.
References
- 13.9 - Example: Visual Motion Area MT
- 2Structure and function of visual area MT
- 3A selective impairment of motion perception following lesions of the middle temporal visual area (MT)
- 4Functional properties of neurons in middle temporal visual area of the macaque monkey. I. Selectivity for stimulus direction, speed, and orientation
- 5Direction and orientation selectivity of neurons in visual area MT of the macaque
- 6The analysis of visual motion: a comparison of neuronal and psychophysical performance
- 7Cortical microstimulation influences perceptual judgements of motion direction
- 8Microstimulation in visual area MT: effects on direction discrimination performance
- 9The Contribution of Area MT to Visual Motion Perception Depends on Training
- 10A motion area in human visual cortex
- 11Selective disturbance of movement vision after bilateral brain damage

Add more perspectives or context around this Post.